IL-8 is an important chemokine involved in CF inflammation. The identification of novel B cepacia and P aeruginosa target candidate antigens that induce host cells to oversecrete IL-8 may lead to new therapeutic approaches.
Cystic fibrosis (CF) is a life-threatening autosomal recessive disorder caused by mutations in the CF transmembrane conductance regulator (CFTR). Although over 1,000 mutations of the CFTR gene have been identified, the most common mutant allele is the _F508 mutation. This mutation is due to a three-nucleotide deletion at the 508th codon that causes the deletion of a phenylalanine residue, ultimately causing defective intracellular processing of the CFTR protein. Because CFTR is an important chloride channel, defects in CFTR result in thickening of the airway surface liquid, which subsequently causes the mucociliary beat to become compromised. Thus, clearance of bacteria is impaired, resulting in increased microbial colonization. In an attempt to combat the overload of pathogens, the host secretes enhanced pro inflammatory cytokines and chemokines. Indeed, marked increases in proinflammatory cytokines and chemokines in CF patients, and, in particular, in the chemokine interleukin 8 (IL-8), have been reported.1 An overproduction of IL-8 leads to an influx of neutrophils, ultimately contributing to the chronic, debilitating lung damage observed in CF patients.2
Common Pathogens
The predominant pathogens that colonize CF patients are Staphylococcus aureus, Haemophilus influenzae, Pseudomonas aeruginosa, and the newly emerging CF pathogen Burkholderia cepacia.
S. aureus is characteristically the first pathogen to colonize the respiratory tract of CF patients. Although S. aureus is typically isolated from the upper respiratory tract, some evidence suggests this pathogen also colonizes the lower airways of CF patients.3 Ratner et al4 have demonstrated that S. aureus potently induces epithelial cells to secrete IL-8. S. aureus secretes a number of extracellular proteins. Chromosomally encoded global regulators such as accessory gene regulator (agr) control these proteins. This staphylococcal global gene regulator is known to enhance the expression of secretory proteins. Thus, S. aureus agr mutants do not induce epithelial cells to secrete IL-8 as potently as the wild type pathogen.4 Although the ligand that induces IL-8 is not known, it is clear that IL-8 induction by S. aureus in CF is agr-dependent.
H. influenzae colonizes the lower airways of 8% of CF patients.3 The majority of H. influenzae isolates from CF patients are noncapsulate and nontypeable and have been associated with chronic bronchitis.5 Recent evidence6,7 demonstrates that H. influenzae can potently induce epithelial cells to secrete IL-8. The major H. influenzae virulence factor for IL-8 induction in respiratory epithelial cells is lipo-oligosaccharide. Several unidentified virulence factors have also been reported.6
P. aeruginosa is the predominant CF pathogen and is commonly isolated from 80% of CF patients. A classical feature of P. aeruginosa strains infecting CF patients is that they mutate into the mucoid, exopolysaccharide alginate-producing variants of the colonizing strain. Conversion of P. aeruginosa to the mucoid phenotype is dependent upon the activation of the alternative stress response sigma factor AlgU. Firoved et al8 have demonstrated that lipoproteins controlled by AlgU might have proinflammatory activity. Lipopeptides from P. aeruginosa were shown to induce human peripheral monocytes to secrete IL-8. Findings from their study suggested that some of the genes coexpressed with the alginate system during conversion to mucoid might play a role in the inflammatory processes of CF. Other factors of P. aeruginosa that induce cells to secrete IL-8 include secretory factors such as pyocyanin9,10; pilin, flagellin, autoinducer4,11,12; lipopolysaccharides13,14; elastase15; phospholipase C16; and nitrite reductase.17 DiMango et al18 have demonstrated that IL-8 induction by P. aeruginosa is mediated via pilin and flagellin. In their work, isogenic mutants lacking pilin and flagellin did not bind to the epithelial cells and did not elicit IL-8 induction, confirming that pilin and flagellin are important IL-8 inducing factors. Furthermore, this group found that an exoproduct secreted by P. aeruginosa during chronic infection also stimulated epithelial cells to secrete IL-8. It is apparent that P. aeruginosa organisms have many virulence factors that can potently induce cells to secrete IL-8. Thus, further investigations of these and other IL-8–inducing factors will facilitate understanding how P. aeruginosa interacts with host cells to induce IL-8 secretion.
B. cepacia is an intrinsically antibiotic-resistant phytopathogen. Originally isolated from soft onion rot, it is increasingly becoming a deadly human pathogen in the CF community. With better culture conditions and improved identification methods, the taxonomy of B. cepacia has now enhanced understanding of the new genus within which it has been placed. At least nine novospecies, termed genomovars, now constitute the B. cepacia complex (BCC).19 B. cepacia organisms from genomovars I through VIII have been isolated from CF patients, but the most pathogenic isolates belong to genomovar III of the BCC, followed by Burkholderia multivorans (formerly genomovar II).20
 |
 |
|
|
||
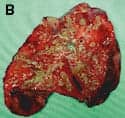
B. cepacia is a highly invasive organism that can migrate across the epithelial barrier of the lung and invade the parenchyma and capillaries.21 In some cases, B. cepacia-infected patients eventually suffer from B. cepacia syndrome, with symptoms that include excessive inflammation, bacteremia, and leukocytosis that result in rapid and fatal clinical deterioration.22 This syndrome is not found in CF patients colonized by any of the other common CF pathogens. The severity of inflammation in B. cepacia-infected human lung is shown in the figure. Panel A shows a normal healthy human lung removed at autopsy. A specimen of a B. cepacia-infected human lung removed from a CF patient during a transplantation procedure at Childrens Hospital, Los Angeles, is shown in panel B. The B. cepacia isolated from this lung was confirmed as belonging to genomovar III of the BCC. In contrast to the normal lung, the B. cepacia-infected CF lung tissue shows heavy infiltration of bloody and yellow exudate. It is clear that this pathology causes lung failure due to severe lung inflammation. It is also evident in this B. cepacia-infected CF lung that inflammation is not localized, but affects the whole lung.
Current Work
The focus of our laboratory at Childrens Hospital Los Angeles, Los Angeles, Calif, is to investigate the mechanism of IL-8 induction by B. cepacia. Our initial studies23 indicate that B. cepacia-mediated IL-8 secretion by human lung epithelial cells occurs via CD14 and p38 mitogen-activated, kinase-dependent pathway. In our more recent studies,24 clinical isolates of the BCC were found to induce human CFTR-mutated lung cells to exaggerate IL-8 secretion excessively, compared with matched CF-corrected cells. Moreover, the highly transmissible clinical isolate of B. cepacia from genomovar III of the BCC induced CF cells to secrete four times more IL-8 than clinical isolates from the other genomovars. At present, we are elucidating which B. cepacia virulence factors may be contributing to this induction of IL-8 by CF cells. Other groups that have investigated IL-8 induction by B cepacia include Palfreyman et al25 and Sajjan et al.26 Their studies have shown that other B. cepacia factors that play a role in inducing IL-8 include secretory proteins and pilin. Thus, further studies need to be conducted to unravel all of the virulence factors that may contribute to B. cepacia-mediated IL-8 induction by epithelial cells.
Conclusion
With better culture conditions and improved identification methods, the identification of CF pathogens continues to increase. Although P. aeruginosa remains the most prevalent CF pathogen, certain strains of B. cepacia are proving highly transmissible between CF patients. Thus, if B. cepacia is uncontrolled, this pathogen may become more prevalent in CF patients. B. cepacia colonizes drinking containers, respiratory equipment, disinfectants, and injection devices.27,28 There is, however, growing speculation that the principal source of B. cepacia infection in CF patients is the respiratory secretions of colonized patients. As a result, the segregation of B. cepacia-infected individuals is currently being practiced. As B. cepacia is resistant to antibiotics, management of B cepacia-infected patients is very challenging. Furthermore, patients infected by isolates from the BCC are excluded from undergoing lung transplantation29 due to increased mortality. CF patients infected with genomovar III of the BCC before transplantation were found to have increased early mortality after transplantation. B. cepacia is proving to be a complex pathogen that contains many virulence factors. As IL-8 is an important chemokine involved in CF inflammation, the identification of novel B. cepacia and P. aeruginosa target candidate antigens that induce host cells to oversecrete IL-8 may lead to new therapeutic approaches.
Krisanavane Reddi, PhD, is Assistant Professor of Research, Surgery, Childrens Hospital Los Angeles Research Institute, Los Angeles, Calif. Stephen B. Phagoo, PhD, is Assistant Professor of Research, Surgery, Childrens Hospital Los Angeles Research Institute, Los Angeles, Calif. Marlyn S. Woo, MD, is Assistant Professor of Clinical Pediatrics, Childrens Hospital Los Angeles Research Institute, Los Angeles, Calif. Kathryn D. Anderson, MD, is Chief of Surgery, Childrens Hospital Los Angeles Research Institute, Los Angeles, Calif. David Warburton, DSc, MD, is Professor of Surgery, Childrens Hospital Los Angeles Research Institute, Los Angeles, Calif.
Acknowledgement
The authors thank Marlyn Woo, MD, for the photographs used here. This work was supported by National Institutes of Health Program Project Grant HL60231 (D.W. and K.R.), the Cystic Fibrosis Foundation (S.B.P.), and a fellowship from the Childrens Hospital, Los Angeles, Research Institute (S.B.P.).
References:
1. Dean TP, Dai Y, Shute JK, Church MK, Warner JO. Interleukin-8 concentrations are elevated in bronchoalveolar lavage, sputum, and sera of children with cystic fibrosis. Pediatr Res. 1993;34:159-161.
2. Dinwiddie R. Pathogenesis of lung disease in cystic fibrosis. Respiration. 2000;67:3-8.
3. Armstrong DS, Grimwood K, Carlin JB, Carzino R, Olinsky A, Phelan PD. Bronchoalveolar lavage or oropharyngeal cultures to identify lower respiratory pathogens in infants with cystic fibrosis. Pediatr Pulmonol. 1996;21:267-275.
4. Ratner AJ, Bryan R, Weber A, et al. Cystic fibrosis pathogens activate Ca2+-dependent mitogen-activated protein kinase signaling pathways in airway epithelial cells. J Biol Chem. 2001;
276:19267-19275.
5. Foxwell AR, Kyd JM, Cripps AW. Nontypeable Haemophilus influenzae: pathogenesis and prevention. Microbiol Mol Biol Rev. 1998;62:294-308.
6. Clemans DL, Bauer RJ, Hanson JA, et al. Induction of proinflammatory cytokines from human respiratory epithelial cells after stimulation by nontypeable Haemophilus influenzae. Infect Immun. 2000;68:4430-4440.
7. Frick AG, Joseph TD, Pang L, Rabe AM, St Geme JW 3rd, Look DC. Haemophilus influenzae stimulates ICAM-1 expression on respiratory epithelial cells. J Immunol. 2000;164:4185-4196.
8. Firoved AM, Boucher JC, Deretic V. Global genomic analysis of AlgU (sigma(E))-dependent promoters (sigmulon) in Pseudomonas aeruginosa and implications for inflammatory processes in cystic fibrosis. J Bacteriol. 2002;184:1057-1064.
9. Denning GM, Wollenweber LA, Railsback MA, Cox CD, Stoll LL, Britigan BE. Pseudomonas pyocyanin increases interleukin-8 expression by human airway epithelial cells. Infect Immun. 1998;66:5777-5784.
10. Leidal KG, Munson KL, Denning GM. Small molecular weight secretory factors from Pseudomonas aeruginosa have opposite effects on IL-8 and RANTES expression by human airway epithelial cells. Am J Respir Cell Mol Biol. 2001;25:186-195.
11. Bryan R, Kube D, Perez A, Davis P, Prince A. Overproduction of the CFTR R domain leads to increased levels of asialoGM1 and increased Pseudomonas aeruginosa binding by epithelial cells. Am J Respir Cell Mol Biol. 1998;19:269-277.
12. DiMango E, Ratner AJ, Bryan R, Tabibi S, Prince A. Activation of NF-kappaB by adherent Pseudomonas aeruginosa in normal and cystic fibrosis respiratory epithelial cells. J Clin Invest. 1998;101:2598-2605.
13. Escotte S, Danel C, Gaillard D, et al. Fluticasone propionate inhibits lipopolysaccharide-induced proinflammatory response in human cystic fibrosis airway grafts. J Pharmacol Exp Ther. 2002;302:1151-1157.
14. Tabary O, Escotte S, Couetil JP, et al. Genistein inhibits constitutive and inducible NFkappaB activation and decreases IL-8 production by human cystic fibrosis bronchial gland cells. Am J Pathol. 1999;155:473-481.
15. Kon Y, Tsukada H, Hasegawa T, et al. The role of Pseudomonas aeruginosa elastase as a potent inflammatory factor in a rat air pouch inflammation model. FEMS Immunol Med Microbiol. 1999;25:313-321.
16. Konig B, Vasil ML, Konig W. Role of haemolytic and non-haemolytic phospholipase C from Pseudomonas aeruginosa in interleukin-8 release from human monocytes. J Med Microbiol. 1997;46:471-478.
17. Sar B, Oishi K, Wada A, Hirayama T, Matsushima K, Nagatake T. Nitrite reductase from Pseudomonas aeruginosa released by antimicrobial agents and complement induces interleukin-8 production in bronchial epithelial cells. Antimicrob Agents Chemother. 1999;43:794-801.
18. DiMango E, Zar HJ, Bryan R, Prince A. Diverse Pseudomonas aeruginosa gene products stimulate respiratory epithelial cells to produce interleukin-8. J Clin Invest. 1995;96:2204-2210.
19. Mahenthiralingham E, Baldwin A, Vandamme P. Burkholderia cepacia complex infection in patients with cystic fibrosis. J Med Microbiol. 2002;51:533-538.
20. De Soyza A, McDowell A, Archer L, et al. Burkholderia cepacia complex genomovars and pulmonary transplantation outcomes in patients with cystic fibrosis. Lancet. 2001;358:1780-1781.
21. Sajjan U, Corey M, Humar A, et al. Immunolocalisation of Burkholderia cepacia in the lungs of cystic fibrosis patients. J Med Microbiol. 2001;50:535-546.
22. Govan JRW, Deretic V. Microbial pathogenesis in cystic fibrosis: mucoid Pseudomonas aeruginosa and Burkholderia cepacia. Microb Rev. 1996;60:539-574.
23. Reddi K, Phagoo SB, Anderson KD, Warburton D. Burkholderia cepacia induced IL-8 gene expression in an alveolar epithelial cell line: signaling through CD14 and MAPK. Pediatr Res. In press.
24. Reddi K, Phagoo SB, Anderson KD, Warburton D. The cystic fibrosis pathogen Burkholderia cepacia, potently induces IL-8 synthesis in CFTR mutated epithelial cells. Am J Respir Crit Care Med. 2002;164: published abstract.
25. Palfreyman RW, Watson ML, Eden C, Smith AW. Induction of biologically active interleukin-8 from lung epithelial cells by Burkholderia (Pseudomonas) cepacia products. Infect Immun. 1997;65:617-622.
26. Sajjan U, Ackerley C, Forstner J. Interaction of cblA/adhesin-positive Burkholderia cepacia with squamous epithelium. Cellular Microbiol. 2002;4:73-76.
27. Cimolai N, Trombley C. Insertional sequence primers for Bordetella pertussis diagnostic polymerase chain reaction differentiate strains of Pseudomonas cepacia. J Infect Dis. 1995;172:293-295.
28. Ouchi K, Abe M, Karita M, Oguri T, Igari J, Nakazawa T. Analysis of strains of Burkholderia (Pseudomonas) cepacia isolated in a nosocomial outbreak by biochemical and genomic typing. J Clin Microbiol. 1995;33:2353-2357.
29. Aris RA, Routh JR, Lipuma JJ, Heath DG, Gilligan PH. Lung transplantation for cystic fibrosis patients with Burkholderia cepacia complex. Am J Respir Crit Care Med. 2001;164:2102-2106.